
You have a Mytilicola intestinalis, Steuer, 1902 – Red worm disease
Synonyms: None known.
Common names: The condition caused by this parasitic copepod is known as “red worm disease”; Le copepod rouge (FR).
This is a parasitic copepod living in the intestine of bivalves, in particular mussels, but also oysters. It has a characteristic red color, which makes it very conspicuous inside the host. Adults have elongate (“worm-like”) bodies with very short appendages. Cephalic appendages, including mouthparts have been described and illustrated (Hockley, 1951). Females are larger than males, 9 mm against 4.5 mm (Gee & Davey, 1986). The paired external egg-sacs of the females are also red. Adult copepods are found in the posterior part of the intestine of the mussel whereas the copepodite stages are found in the stomach digestive gland, and anterior part of the intestine (Gresty, 1992).
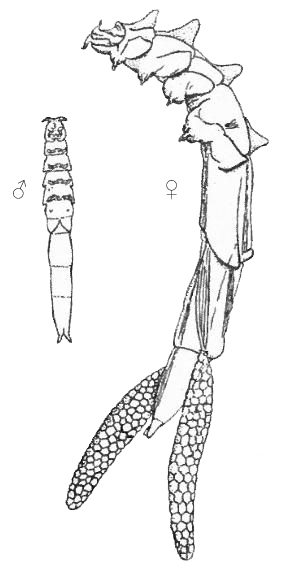
Mytilicola intestinalis, male and ovigerous female (from Hockley 1951)
Similar species: Mytilicola orientalis, 1935 (syn: Mytilicola ostreae, Wilson, 1938), NON Myicola ostreae (Hoshina & Sugiura, 1953) (=Mytilicola ostreae Hoshina & Suguira, 1953). The latter species parasitizes gills rather than intestine of bivalves. Both species are found in European countries, often associated with the introduced Pacific oyster, Crassostrea gigas (Thunberg, 1793). More species of similar parasitic copepods from mussels and oysters have been described from the Pacific Ocean.
Native area: Most likely the Mediterranean where it was first described from Mytilus galloprovincialis Lamarck, 1818 in the Adriatic Sea. In 1914 it was found in Mytilus edulis Linnaeus, 1758 in the Mediterranean (Bolster, 1954).
Introduced area: In 1937 a single specimen was found in Mytilus edulis at Portsmouth in southern England, but it was not until 1951 that this was published, and 1947 is usually listed as the first date (Campbell, 1970). In 1938 it was found in the German Wadden Sea (Bolster, 1954). M. intestinalis was first found in the Limfjord in 1964 (Theisen, 1964, 1966) and only in 1994 were the first infected mussels found in the Danish Wadden Sea (Theisen, pers comm.). The first record from the French Atlantic coast was from 1949 (Goulletquer et al., 2002). In the Netherlands the first observations were also from 1949 (Wolff, 2005), and in Belgium from 1950 (Kerckhof et al., 2007). In Ireland the first record was from 1948 (Minchin, 2007). It has not yet been found in Norway or Sweden (Hopkins, 2001; Främmande arter, alert list). Nor has it been found in Poland, Lithuania, Latvia, Estonia or Finland.
There is an old record of this species from the Malacca Strait, Indian Ocean (Wickstead, 1960), but there are no records of the species since then, so this record must be considered dubious.
Vector: Probably infected mussels transported as fouling on ships’ hulls or infected mussels transferred for aquaculture (Hockley, 1951; Theisen, 1966; Korringa, 1968). Secondary dispersal through free swimming larvae seems unlikely as the actively swimming stages last only a few days (Hockley, 1951).
Most of the information available is on prevalence and effects of the parasite on the host animals, and very little information is available about the ecology of the parasite. Mytilicola intestinalis in Mytilus galloprovincialis seems fairly rare in the Aegean Sea, infecting about 11% of the mussels with 1-4 individuals per mussel (Rayyan et al., 2004). However, these were mussels cultured on hanging ropes, which are less prone to infection by this parasite (Theisen, 1987). It also seems rare in M. galloprovincialis from Italy whereas almost 60% of the M. galloprovincialis imported to Italy from Spain were infected (Trotti et al., 1998), and even higher infection rates were recorded from cultured mussels from the Spanish Atlantic coast (Fuentes et al., 1995).
The effect on Mytilus edulis depends on the number of parasites, but even the presence of one copepod may cause visible effects (Korringa, 1968), but see also below for impacts. The highest number of parasites are found in large sized mussels. This has been attributed to higher filtration rates rather than age (Williams, 1967; Paul, 1983), though this has also been contested (Davey, 1989). In most places relatively low numbers (1-10 individuals) of parasites are found (Grainger, 1951; Williams, 1969; Theisen, 1987). Mussels in beds on the bottom are more heavily infected than mussels on vertical surfaces away from the bottom (Korringa, 1968). The smallest mussel infected was about 10 mm long (Williams, 1967). Fewer parasitic copepods are found in mussels infected by the shell-boring polychaete Polydora ciliata (Johnston, 1838) (Williams, 1968). Adult females have almost the same diameter as the intestine of the host, but a groove formed by appendages on the dorsal side permits the flow of food to pass, the copepod only picking up enough for its own nutrition (Hockley, 1951). What constitutes the diet of M. intestinalis has been discussed, but it seems certain that it does not include mussel tissues (Davey, 1989; Gresty, 1992). If an adult parasite is expelled it is not able to move enough to find a new host (Grainger, 1951).
Mytilicola intestinalis is able to tolerate a wide range of temperatures, at least between -1.4° and 30° C, and also is extremely euryhaline (Korringa, 1968).
Reproduction: Fertilization is assumed to take place within the host (Grainger, 1951), and sex ratio is always skewed towards males (Hockley, 1951; Theisen, 1966). It has been claimed that it needs at least 18° C to initiate reproduction (Williams, 1969), but may continue as long as the temperature remains above 6° C (Bolster, 1954), though this has been contested (Koringa, 1968; Davey, 1989). There seems to be two annual spawning periods (Williams, 1969; Davey et al., 1978), but in northern European waters the winter spawning (December – February) may not be successful in infecting new hosts (Williams, 1969). Mytilicola intestinalis produces 200-300 eggs (Thieltges et al., 2008). Eggs detached and grown in the laboratory hatched after 7 days at 18° C (Hockley, 1951). There are a nauplius and a metanauplius larval stages, which are free-swimming. The infective stage is the first copepodite, and there are 5 successive copepodite stages before it reaches the immature adult stage (Gee & Davey, 1986). The nauplius is positively phototropic. Some copepodite I specimens have positive phototaxis, others negative (Grainger, 1951), and this stage is only free swimming for 3-4 days after which it moves to the bottom to search for a host (Campbell, 1970). Development from release of nauplius to copepodite I is about 2 days at 13-14° C. The free swimming larvae have poorly developed mouth parts and may not need to feed. The copepodite II is reached after about 12 days and copepodite III after 15-20 days (Grainger, 1951), though the infective copepodite I may survive longer if a host is not found (Gee & Davey, 1986).
The physical impact to mussel tissues is caused by the copepod moving about and scraping the intestinal epithelium and scar tissue formation (Moore et al., 1977). However, there is considerable controversy about the seriousness of this parasite. Coincidence of outbreaks of M. intestinalis and mass mortality in mussel beds have caused some authors to blame the parasite for the economic losses (Korringa, 1968; Blateau et al., 1992). Other studies have shown little or no effects on the meat content (“condition”) of the mussels (Dethlefsen, 1975; Paul, 1983; Davey, 1989). Some studies have found that high numbers M. intestinalis caused decreased condition of mussels, but only under certain circumstances (Gee et al., 1977). Densities of more than 10 copepods per mussel may render the mussels unmarketable due to low condition. Apparently only mussels close to or on the bottom are infected (Korringa, 1968; Theisen, 1987). Hence long-line culture should not be affected, as long as spat is not transferred from infected mussel beds. Low rates of infestation have been found in line cultures in German waters (Buck et al. 2005) and high rates of infection has been found in Spain (Paul, 1983; Fuentes et al., 1995), where part of the seed mussels came from intertidal beds. Other effects on the mussel host include retarded spawning, reduced shell growth, decreased filtration rate, and weakened byssus attachment, although most of these serious effects were only found in early studies and could not be confirmed in more recent studies (Bayne et al., 1978; Davey, 1989). Whether this is due to inappropriate procedures or faulty observations in the early studies, or whether the mussels have actually adapted to the parasite over time is unknown (Gresty, 1992).
M. intestinalis may also infect oysters, and the native European flat oyster, Ostrea edulis Linnaeus, 1758, seems to be more susceptible to infection than Pacific oyster, Crassostrea gigas (Dare, 1982), and although they are not heavily infected, transport of live seed oysters may transfer the parasite to new areas.
Mytilicola intestinalis has been declared a “controlled pest” in the U.K., which means that transport of mussels from areas infected is carefully monitored (Gresty, 1992). Decreasing stocking density in mussel farms may decrease the problem (Blateau et al., 1992), and in France mussel farmers have created a union which works for voluntary decreasing stocking density to prevent outbreaks of this parasite (Mongruel & Thébaud, 2006). Also, various pesticides have been tested as curative treatment for this parasite (Blateau et al., 1992), but this is highly toxic to a number of aquatic organisms. Free swimming stages can be killed by chlorine (Korringa, 1968). Probably the most efficient measure is control of transfer of live mussels from infected areas.